




















 В разлуке
В разлукеРазговор c оставшимся
Мария Карпенко поговорила с человеком, который принципиально остается в России: о том, что это ему дает и каких жертв требует взамен
28 ноября 202493574 Была ли крокоутка?
Была ли крокоутка?
 Чем занимаются генетики?
Чем занимаются генетики?
 Кто на Луне главный?
Кто на Луне главный?
 Как возникает лжеистория?
Как возникает лжеистория?
 Что делают медики на войне?
Что делают медики на войне?
 Как рассказать о блокаде Ленинграда?
Как рассказать о блокаде Ленинграда?
 Что придумал Толкин?
Что придумал Толкин?
 Как возникла Вселенная и что с ней будет дальше?
Как возникла Вселенная и что с ней будет дальше?
 Как «Радио Свобода» сохранило запрещенную литературу
Как «Радио Свобода» сохранило запрещенную литературу
 Как спорт превратился в шоу-бизнес?
Как спорт превратился в шоу-бизнес?
 Почему мы такие умные?
Почему мы такие умные?
 Роботы и музыка
Роботы и музыка
 © ТВ Центр
© ТВ ЦентрВ Лектории Политехнического музея лауреат премии «Просветитель», доктор биологических наук и ведущий научный сотрудник Палеонтологического института РАН Александр Марков рассказал об эволюционном пути, который прошли морские ежи и усачи из эфиопского озера Тана, а также о том, что такое переходные формы и почему их важно изучать.
Что такое переходные формы? Вот, к примеру, широко известное в узких кругах животное, которое называется «крокоутка». И это такой символ неверного понимания того, что такое переходные формы. Была история о том, как один американский креационист, выступая по телевизору, показал картинку с крокоуткой и заявил, что поскольку в природе нет такого животного, то эволюция — это ложь. Преподносилось все так: если утки произошли от крокодилов, то пусть эволюционисты покажут нам крокоутку. То есть, по-видимому, креационисты думают, что из теории эволюции следует, будто должны существовать в природе переходные формы, промежуточные между двумя современными видами. Так ли это? Вообще в каком-то смысле — да. Но нужно понимать, что это означает на самом деле, что мы можем найти в качестве переходных форм.

Перед вами эволюционное древо. Родственные связи между организмами — по крайней мере, высшими, в эволюции которых горизонтальный перенос генов играет не очень важную роль. Эволюция более-менее адекватно представляется в виде генеалогического древа, на концах веточек которого сидят современные виды. Вот, скажем, черепаха и птичка. Птичка здесь символизирует всех птиц, а черепаха — всех рептилий. Соответственно ветви символизируют ход эволюции, которая предсказывает, что у двух любых современных видов существовали в прошлом общие предки, а может, существуют и по сей день их мало изменившиеся потомки. Но вообще, если строго говорить о том, что такое переходная форма, то, конечно, это уже вымершее существо. Потому что никакое ныне живущее существо не может быть общим предком каких-то двух других тоже ныне живущих видов.
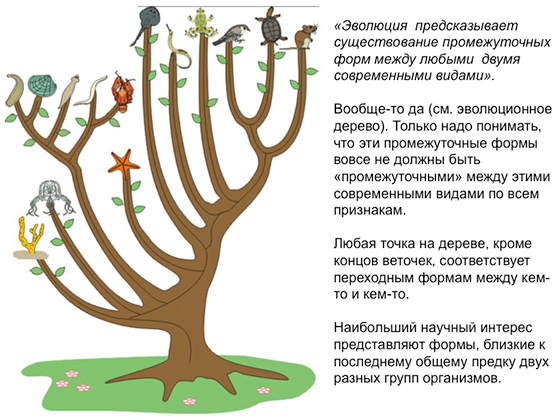
Переходные формы должны были в прошлом существовать между двумя любыми современными видами, но они, конечно, были не обязаны быть промежуточными между этими двумя видами — по всем признакам. В принципе, любая точка на этом дереве — кроме самых концов веточек — соответствует какой-либо переходной форме. Но наибольший научный интерес представляют формы, близкие к последнему общему предку каких-то двух современных групп, то есть близкие к местам разветвления.
В реальной летописи у нас мало шансов найти ископаемый организм, который находился строго на ветви, ведущей к какому-то конкретному виду, то есть абсолютно прямого предка. И вряд ли мы его узнаем, когда найдем, а если и найдем, то вряд ли докажем, что это он. Как правило, мы находим представителей каких-то маленьких боковых веточек, ответвлений, что-то похожее, что-то близкое, какого-то из вымерших родственников.
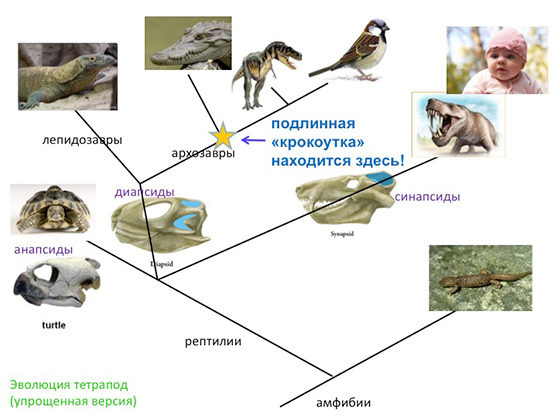
А что касается крокоутки, то — что на самом деле мы должны ожидать увидеть в этой ископаемой летописи на месте крокоутки? Значит, вот это очень грубая, примитивная, упрощенная схема эволюционного древа тетраподов (надкласс позвоночных, четвероногие, перешедшие к жизни на суше и в воздушной среде. — Ред.). Вот их общий предок — когда рыбы вышли на сушу, так сказать. Первое разветвление — одна ветвь ведет к современным амфибиям, другая — ко всем остальным. Рептилии делятся на три ветви, взаимоотношения между которыми спорны, но мы сейчас не об этом. Самые большие ветки — это диапсиды с двумя дырками в черепе для прохода жевательных мышц и синапсиды с одной дыркой. Синапсиды — это наши прямые предки. Между первыми синапсидами и млекопитающими есть ряд замечательных промежуточных форм — зверозубые рептилии так называемые. Диапсиды, в свою очередь, разделились где-то в пермском, а может, даже и в каменноугольном периоде на лепидозавров, к которым относятся ящерицы, змеи и вся их родня, и на архозавров, к которым относятся крокодилы и динозавры. Птицы тоже относятся к архозаврам, так как их ближайшей родней являются те же динозавры. Соответственно аналог крокоутки находится на самом деле где-то здесь, и его правильно называть не крокоуткой, а «последним общим предком всех современных архозавров». Палеонтологи нашли несколько форм подходящего возраста, подходящего строения, то есть базальных архозавров — форм, близких к последнему предку архозавров. Вот одна из этих форм, которая называется «архозавр россикус», из самого конца пермского периода, описанного нашим академиком Леонидом Петровичем Татариновым.

Вот как примерно выглядел последний общий предок всех птиц и крокодилов. Вот еще ряд реконструкций базальных архозавров. Все они более или менее были близки к той самой форме, которая и была последним общим предком крокодилов и птиц. Кто из них конкретно был ближе всего к этому предку, мы не знаем и вряд ли уж в таких деталях когда-нибудь разберемся. Но мы точно знаем, что было животное примерно вот такого плана строения — базальный архозавр, и от него затем эволюционировали, с одной стороны, крокодилы, с другой — произошли первые динозавры в конце триасового периода, которые потом подразделялись на ящеротазовых и птицетазовых, и одна из ветвей ящеротазовых динозавров дала начало птицам. Вот что было на самом деле вместо крокоутки.
Теперь разберем отдельные примеры — начнем с камбал. Вопрос о происхождении камбал представлялся долгое время одним из самых трудных в теории эволюции. Потому что теория говорит о том, что организм эволюционирует медленно и постепенно, путем накопления мелких изменений (и каждое последующее изменение должно быть чем-то полезно, должно поддерживаться отбором, должно приносить какую-то пользу, чтобы организмы с этим новым признаком могли оставлять больше потомства, чем организмы без него). И соответственно многим казалось — и прежде всего креационисты на этом настаивали, — что невозможно представить себе существование переходных форм между нормальной рыбой и камбалой.

Камбалы, как известно, лежат на боку на дне, и оба их глаза находятся на одной стороне головы. Понятно, что в таком законченном виде данная адаптация им очень удобна: лежать, притаившись, и оба глаза смотрят наверх, откуда может появиться добыча. Было бы совсем не так удобно, если бы один глаз был на другой стороне головы и смотрел в песок. Начальные этапы процесса перемещения глаза с одной стороны головы на другую не могли приносить никакой пользы — глаз ведь не мог сразу взять и перепрыгнуть. Он должен был, скорее всего, двигаться постепенно, из поколения в поколение, сдвигаясь на миллиметр в тысячу лет. И совершенно непонятно, какую пользу могли получить предки камбалы из того, что их нижний глаз — тот, который утыкается в песок, — немножко сместится вбок. Вот в чем была загвоздка. И креационисты говорили: вот пример адаптации, которая не может развиться постепенным путем, потому что переходные формы не могут существовать, не могут быть поддержаны отбором.
Казалось бы, такой убедительный аргумент. Но его удалось опровергнуть в 2008 году, когда палеонтологи нашли рыб, которые вопреки всем этим представлениям как раз демонстрируют переходную морфологию. И вот это тоже надо пояснить. Камбала появляется на свет с нормальной головой. В ходе индивидуального развития камбалы один из ее глаз совершает удивительное путешествие. У маленького малька оба глаза расположены как у нормальных рыб — один слева, другой справа, но затем в ходе развития рыбки один глаз начинает смещаться и оказывается сначала на макушке, а потом переползает полностью на другую сторону, и в результате оба глаза оказываются рядом. Эта история повторяется, проигрывается заново при рождении каждой камбалы. Это пример того, как какие-то эволюционные изменения рекапитулируются, повторяются в индивидуальном развитии.

Как могли бы выглядеть промежуточные формы камбалы? Есть такой псеттод. Примитивная камбалообразная рыба, которая отличается от остальных, более продвинутых, камбал тем, что у нее этот глаз находится на ребре, на верней стороне головы, а не на боку, то есть находится в таком промежуточном состоянии. Это не реальный предок настоящих камбал, а современная рыба, которая показывает нам, как могли быть устроены предки. Но все равно у псеттода глаз что-то уже видит, когда он лежит на боку, он уже как бы полезен. А как быть с переходными формами, у которых второй глаз еще внизу? И вот как раз такие ископаемые рыбы и были обнаружены.
Два рода ископаемых рыб — Amphistium и Heteronectes, у которых глаза (именно у взрослых рыб) как раз находятся в промежуточном состоянии. Были обнаружены и левые, и правые формы, то есть у одних к макушке смещен левый глаз, у других правый. Собственно, таким вот образом палеонтологи доказали, что промежуточные формы между нормальными рыбами и камбалами возможны. Они точно были — вот они. Другое дело, как ответить на вопрос: а зачем они были? Зачем этим рыбам нужен был вот такой смещенный глаз, если он все равно смотрел в песок? И ответить, полагаясь на ископаемый материал, очень трудно, потому что мы не знаем, как вели себя эти рыбы. Но наблюдения за современными камбалообразными дали возможность высказать правдоподобную гипотезу. Дело в том, что некоторые камбалообразные поднимают тело над поверхностью дна, используя плавники. И промежуточные формы — между нормальными рыбами и камбалами — могли таким способом осматривать своим смещенным глазом дно. Они могли приподниматься на плавниках и смотреть, не ползает ли что-то вкусненькое по поверхности дна (второй глаз при этом смотрел вверх). Для такого выглядывания смещенный глаз гораздо удобнее, чем тот, что находится на своей изначальной позиции, на боку.

Дальше я хотел вам рассказать про морских ежей, которыми я изначально занимался как палеонтолог. Обычно я на своих лекциях излагаю то, что в книжках прочел, но сегодня решил сделать исключение и рассказать кое-что о том, чем я сам, своими руками, занимался как ученый. В палеонтологии морских ежей есть масса интереснейших переходных форм. Правда, чтобы понять, что они переходные и что они интересные, нужно достаточно детально с ними познакомиться. На примере морских ежей мы видим, что иногда, когда палеонтологическая летопись достаточно подробна (а для морских ежей она именно такая, потому что они хорошо сохраняются в ископаемом состоянии, особенно те из них, что зарывались в грунт, вели, так сказать, закапывающийся образ жизни), известно много хорошо изученных, подробных эволюционных рядов. И в последовательности слоев — чем глубже слой, тем древнее останки — мы встречаем постепенно меняющихся морских ежей.
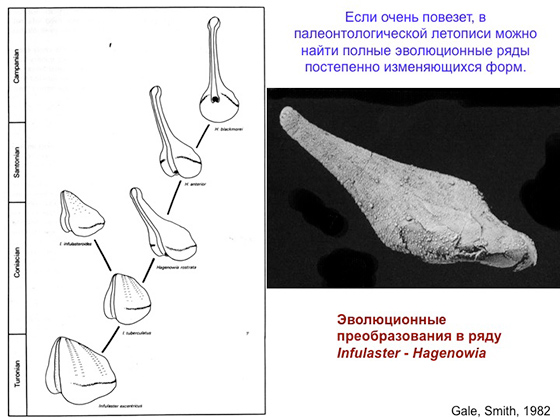
Вот здесь показан эволюционный ряд необычных морских ежей Infulaster — Hagenowia. Они закапывались в грунт. Однако и закапывающемуся ежу нужно иметь доступ к поверхности, чтобы в его нору поступала свежая вода и падал осадок со дна, которым он питается. Многие другие закапывающиеся морские ежи решали эту проблему, но вот именно эта группа придумала кое-что необычное. Она в итоге отрастила себе такой вырост на панцире, который торчал на поверхности, пока сам еж сидел в грунте.
Вот пример эволюционного ряда, очень подробно изученный. В левом столбце вы можете прочитать: верхний турон, коньяк, сантон, нижний кампан, верхний кампан. Это все подразделения верхнего мела — временные интервалы. Каждая пара картинок соответствует одному хроновиду, то есть видам, которые сменяли друг друга во времени. И здесь же показаны морфологические изменения от слоя к слою, от самых древних форм к самым молодым. Например, рот изначально был здесь, а потом постепенно смещался вверх, ближе к переднему краю панциря, пока не оказался совсем впереди. И это морфологический признак, который менялся направленно. Такой ход эволюции был связан в данном случае с тем, что ежи приспосабливались к закапыванию все в более тонкозернистый осадок. С переходными формами здесь тоже все понятно: каждый вид является переходным между предыдущим и будущим видами. И это редкость и большая удача, когда полный эволюционный ряд удается найти разложенным по слоям разного возраста — как на картинке. Но бывают гораздо более сложно расшифровываемые случаи. И сейчас я расскажу о переходных формах, которые были в эволюции так называемых неправильных морских ежей.

Вообще класс морских ежей делится на два подкласса — правильные и неправильные, и мы поговорим сейчас о неправильных. Вот это милое пушистенькое не пойми что и есть типичный представитель неправильных ежей Echinocardium cordatum. Но это у него не шерстка и не пух, а тоненькие известковые иголочки, какие и положено иметь морскому ежу. Echinocardium — закапывающееся существо. На картинке он весь в иголочках, но если ему их удалить, то останется панцирь. И чтобы вы понимали, о чем я дальше буду говорить, мне придется познакомить вас со строением морских ежей и терминами, без которых совершенно невозможно будет ничего рассказать… Морские ежи вообще мало известны широкой публике, так что знакомство это будет очень кстати.
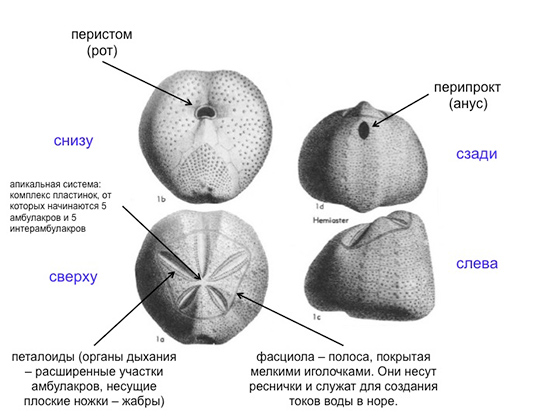
Вот панцирь типичного неправильного морского ежа. Вид сверху, вид снизу, вид сзади и сбоку. На картинке все подписано. Панцирь очищен от иголок и эпителия, то есть перед нами фактически скелет морского ежа. Снизу и спереди у него рот — и это отверстие в панцире называется перистомом. На задней стороне панциря у него перипрокт, в котором находится анальное отверстие. Вид сверху: передняя борозда, пищесобирающая структура, там амбулакральные длинные ножки сидят, которые строят вертикальный канал в грунте. Четыре лепесточка, как у цветка, называются петалоидами — это органы дыхания. Расширенные участки рядов пластинок с ножками, которые называются амбулакрами. Вообще у морских ежей пятилучевая симметрия, и их панцирь состоит из пяти двойных рядов амбулакральных пластинок, в них есть дырочки, из которых ножки амбулакральные торчат. Пять амбулакров и пять двойных рядов пластинок между амбулакрами — это называется интерамбулакрами. Передний амбулакр у неправильных ежей образует переднюю борозду, остальные четыре — так называемые петалоиды, где ножки превращаются в жабры, через которые гонится вода, поступающая в нору, где еж сидит. Вода омывает жабры — еж дышит. Часто на панцире у неправильных зарывающихся морских ежей есть полоски, обычно образующие замкнутые контуры и покрытые очень мелкими иголочками. Иголочки, в свою очередь, покрыты ресничным эпителием, который создает токи воды, омывающей панцирь ежа. Полоски эти называются фасциолами и служат для вентиляции норы. Еж сидит в норе, и ему нужно, чтобы его омывала всегда свежая вода, чтобы он мог дышать, ему нужно удалять из норы продукты его жизнедеятельности, и необходимо, чтобы вода все время протекала через его нору. И вот эти токи, так необходимые ежу, создаются именно фасциолами. Вот, собственно, и все, что нужно знать о морфологии морских ежей.

Неправильные морские ежи появились в юрском периоде и произошли от более древних правильных ежей. Развитие морского ежа начинается с планктонной личинки, которая называется эхиноплутеусом: из нее в результате метаморфоза получается маленький морской ежик. Он всегда получается сначала правильным, то есть радиально симметричным. Будь то молодой неправильный еж или молодой правильный, крошечный ежик (порядка миллиметра или даже полумиллиметра в диаметре) всегда будет иметь правильную пятилучевую симметрию. Но потом развитие разводит их в разные стороны. У правильного ежа сохраняется пятилучевая симметрия, а у неправильного в процессе роста пятилучевая симметрия превращается в двухстороннюю, то есть у него неравномерно развиваются разные стороны панциря. Дальше перистом смещается вперед. Но самое интересное происходит на верхней стороне: анус находится почти в центре, как у правильных ежей, но потом начинает смещаться назад. И, наконец, у взрослого неправильного ежа — с уже развитыми петалоидами и с передней бороздой — анальное отверстие уходит на задний конец тела, и его сверху не видно.
С какой проблемой столкнулись неправильные морские ежи? Их появление было связано с освоением нового типа питания. Правильные ежи — это животные, которые живут на поверхности морского дна, они более-менее всеядны, у них есть грызущий ротовой аппарат (так называемый аристотелев фонарь) с пятью очень прочными зубами, которыми они все грызут, с чего-то соскребают. Иными словами, они могут питаться водорослями, разными животными, которыми обрастают камни и другие предметы на дне, могут съесть и кого-то мертвого, если найдут. То есть это всеядные формы, активно выбирающие, что им кушать, и их анальное отверстие находится на самой вершине панциря, что им совершенно не мешает.
Неправильные морские ежи — и их появление, я напомню, как раз было связано с другим типом питания — относятся к детритофагам. Они начали питаться просто-напросто грунтом — будь то песок или ил на дне. Они заглатывают грунт в больших количествах и насыщаются той органикой, которая в нем есть. Конечно, это очень низкокалорийная диета. Приходится заглатывать очень много грунта, чтобы получить достаточное количество калорий. Соответственно и отходов много. И если бы у ежа-детритофага анальное отверстие осталось на вершине панциря, то все отходы падали бы на него обратно и он бы повторно заглатывал материал, который уже прошел через его кишечник. Для грунтоеда, как мы понимаем, этот вариант неприемлем. Поэтому первое, что происходит с ежом, переходящим на грунтовую диету, — смещение анального отверстия из центрального положения на бок. Перипрокт должен сместиться из центра куда-то назад. Но как этого добиться? У всех морских ежей с незапамятных времен и по сей день развитие так устроено, что в результате метаморфоза из личинки появляется ежик, у которого перипрокт находится посредине панциря, окруженный пластинками апикальной системы. И оказалось, что это не так-то просто изменить. Очевидно, что древняя связь перипрокта с апикальной системой очень прочная. Апикальную систему нельзя передвинуть, потому что это не просто какие-то там десять пластиночек, а зона роста всего панциря. Именно вокруг апикальной системы формируются новые панцирные пластины. В итоге нужно было оставить апикальную систему наверху, а перипрокт убрать куда-нибудь подальше. На первых порах эволюция неправильных морских ежей выглядела очень причудливо: в разных эволюционных линиях параллельно предпринимались разные неуклюжие попытки сдвинуть перипрокт назад, не убирая апикальную систему. Большинство вариантов оказались тупиковыми и вымерли. Но некоторые оказались успешными, и от них произошли все современные неправильные морские ежи.
Как реально происходит открытие каких-то новых переходных форм? Как палеонтологи это делают? Чтобы объяснить, я расскажу о самой интересной и важной переходной форме, которую мы с моим учителем Андреем Николаевичем Соловьевым открыли и описали, — это переходная форма между двумя семействами неправильных морских ежей. Обычно — и часто так бывает — переходная форма давно найдена. Она лежит в коллекции с этикеточкой, на которой все написано, но при этом никто не догадывается, что это переходная форма между какими-то двумя группами. Чтобы это понять, нужно правильно разобраться в ее развитии, в ее строении и указать то место на эволюционном древе, где она находится.
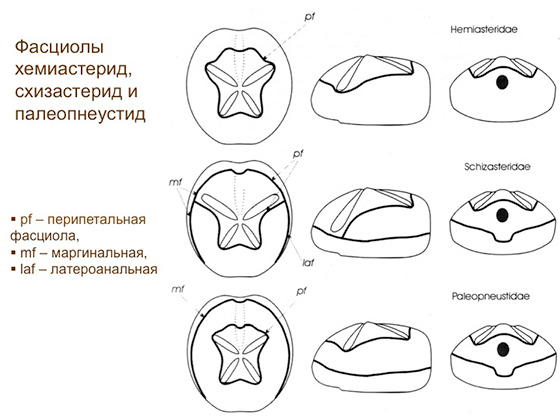
В систематике той группы морских ежей, о которой я сейчас буду говорить, ключевую роль играют фасциолы. Я напомню, что это такое: полосочки на панцире, покрытые мелкими иголочками с ресничным эпителием, которые гонят воду, вентилируют нору. Так вот, разные типы фасциол в отряде Spatangoida появлялись по очереди в ходе эволюции: в среднем меле появляется вот такая перипетальная фасциола, потом в начале верхнего мела, в сеноманском веке, появляются формы, у которых есть какой-то довесок, называемый латероанальной фасциолой, потом идут формы с субанальной фасциолой и так далее. Известно время появления каждого типа фасциол. Три семейства, которыми, собственно, я занимался и о которых далее пойдет речь, называются хемиастеридами, схизастеридами и палеопнеустидами. Отличаются они как раз по фасциолам. У хемиастерид одна фасциола — перипетальная. У схизастерид, как прежде думали, две фасциолы: одна перипетальная, а к ней приделана сзади таким полукольцом фасциола латероанальная. И у третьего семейства — палеопнеустиды — есть перипетальная фасциола, а вокруг панциря идет маргинальная фасциола. Раньше думали, что эволюционно все три семейства связаны следующим образом: предковая система — это хемиастериды с одной перипетальной фасциолой. От них происходят в одну сторону схизастериды, в другую — палеопнеустиды. Думали, что эволюция вот так шла. Казалось, все логично. Но со временем стало очевидно, что вся эта интерпретация неверна, потому что авторы, которые ее разрабатывали, не учитывали (возможно, не знали), как идет развитие морских ежей. На самом деле, если посмотреть, как развивается маленький морской ежик, то сразу все становится понятно. У схизастерид — у них перипетальная фасциола, а к ней сбоку приделана латероанальная — в развитии закладывается маргинальная фасциола. Она закладывается вокруг апикальной системы, а потом вокруг апикальной системы закладываются новые пластинки панциря. По мере роста панциря перипрокт выходит из апикальной системы и перемещается назад, и когда перипрокт выходит, закладывается полукольцом вот такая вещь — задняя часть того, что раньше принимали за перипетальную фасциолу. На самом деле у схизастерид не перипетальная плюс латероанальная, а маргинальная плюс неполная перипетальная, представленная только задней половинкой. И чтобы это понять, нужно было разобраться в индивидуальном развитии морских ежей.
В общем, мы опубликовали нашу работу, в которой один из важнейших выводов говорил о том, что палеопнеустиды произошли от схизастерид, что это очень близкородственные семейства, и это противоречило общепринятым на то время взглядам. А спустя несколько лет английские коллеги выделили ДНК у современных представителей этих семейств и уже по ДНК определили родство. У них получилось то же, что и у нас. То есть наша точка зрения подтвердилась. На что коллеги и сослались, написав, что вот русские ученые Марков и Соловьев несколько лет назад уже высказывали такую точку зрения и вот она и у нас подтвердилась. После чего, когда мы с Андреем Николаевичем обсуждали это, он сказал: интересно, а зачем нужен был этот дурацкий анализ ДНК, мы и без него все по морфологии определили. А я тогда не сказал, но подумал, что молодые современные биологи сказали бы наоборот: кому вообще нужна эта ваша морфология и вся эта палеонтология, если есть такой простой и удобный анализ ДНК. А мне кажется, это хорошо, что есть разные способы, разные науки — как этот пример с палеонтологией и морфологией, с одной стороны, и молекулярной генетикой — с другой.

Мы с вами погрузились достаточно глубоко в такую настоящую, кондовую палеонтологию, где действительно идет поиск настоящих переходных форм между двумя семействами. И следующий сюжет — он же будет последним, потому что остается мало времени, — посвящен проблеме поиска переходных форм в ситуации, когда происходит адаптивная радиация, то есть очень быстрое расхождение из одной предковой формы разных потомков, которые далее эволюционируют абсолютно самостоятельно. Примеры событий адаптивной радиации достаточно часты в эволюции. Например, такое происходит после формирования большого нового озера, когда туда попадают первые случайные рыбешки — и от них целые пучки видов образуются в течение десятков тысяч лет. Адаптивные радиации могут происходить после появления какого-то важного эволюционного новшества или резких изменений условий среды, которые открывают новые возможности. Самый яркий пример адаптивной радиации в истории Земли — это Кембрийский взрыв, о котором вы, наверное, слышали. Еще один замечательный пример — это рыбы-усачи, или барбусы, из эфиопского озера Тана, которое образовалось всего тридцать тысяч лет назад в результате тектонической подвижки, когда возник водопад на Голубом Ниле и отделил новообразовавшееся озеро от бассейна реки. В озеро как-то попали обычные усачи привычного для тех краев вида, и в условиях изоляции за тридцать тысяч лет от одной предковой формы развилось целых 14 форм разных рыб — разных и по внешнему виду, и по поведению, и по питанию, и по образу жизни. Многие из них даже стали хищниками, хотя это семейство карповых, для которого хищничество нехарактерно, — однако в условиях отсутствия конкурентов в лице других хищников они смогли освоить и такое поведение. 14 разных форм! И ихтиологи долго спорили, виды это или разновидности. Вообще это типичная ситуация для ученого сообщества, когда одни — в частности, российские — ихтиологи считали, что это разновидности и не надо называть их полностью сформированными отдельными видами. А голландские ихтиологи взяли и расписали эти 14 форм как 14 отдельных видов, дав им латинские названия. Вообще это так сильно, престижно и круто — взять и одним махом открыть целых 14 новых видов современных рыб. Но правда в том, что рыбы эти еще не утратили способности скрещиваться друг с другом — и прекрасно это делают в лабораторных условиях. Хотя в природе, в реальном озере Тана, они до недавних пор — пока там не началась экологическая катастрофа — почти не скрещивались. На это указывает генетический анализ. До начала усиленного рыболовства это были 14 фактически сформированных видов, и по своему строению челюстей, чешуи, жаберной крышки, плавников многие из них заслуживают родового статуса. И один из наших крупнейших и виднейших ихтиологов абсолютно верно сказал, что если бы эти 14 морфотипов нашли не зоологи, а палеонтологи — уже в виде ископаемых скелетиков, — то их бы описали не только как отдельные виды, но и как отдельные роды, потому что морфологические различия между ними очень серьезные. И вот вопрос: где в данном случае искать переходные формы? Ведь дальше что могло с ними быть? При благоприятных условиях от каждого из этих 14 морфотипов могли бы произойти большие разнообразные группы потомков. И через несколько миллионов лет мы могли бы иметь уже 14 новых отрядов рыб. Где переходные формы в данном случае? Ответ очень простой — потому что мы более-менее знаем, от кого они произошли.
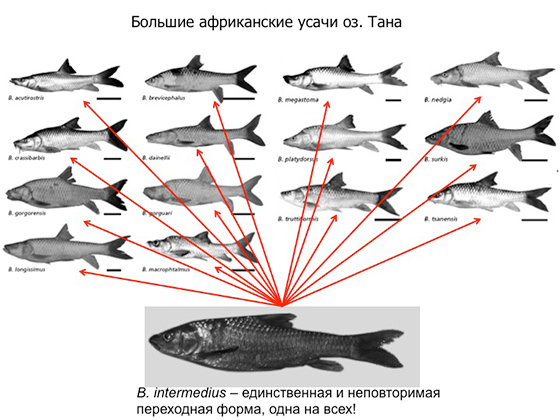
От обычного барбуса, или Barbus intermedius. Именно он является для любой парочки этих морфотипов единственной и неповторимой переходной формой. В палеонтологии всегда нужно учитывать этот фактор: переходные формы могут выглядеть совсем не так, как мы ожидаем. У нас изначально могут быть неточные представления о том, как шла эволюция. И к этому просто нужно быть готовым.
Записала Наталья Кострова
 Поцелуй Санта-Клауса
Поцелуй Санта-Клауса
Запрещенный рождественский хит и другие праздничные песни в специальном тесте и плейлисте COLTA.RU
11 марта 2022
14:52COLTA.RU заблокирована в России
3 марта 2022
14:53Из фонда V-A-C уходит художественный директор Франческо Манакорда
12:33Уволился замдиректора Пушкинского музея
11:29Принято решение о ликвидации «Эха Москвы»
2 марта 2022
18:26«Фабрика» предоставит площадку оставшимся без работы художникам и кураторам
Все новостиВ разлукеМария Карпенко поговорила с человеком, который принципиально остается в России: о том, что это ему дает и каких жертв требует взамен
28 ноября 202493574 В разлуке
В разлукеПроект «В разлуке» начинает серию портретов больших городов, которые стали хабами для новой эмиграции. Первый разговор — о русском Тбилиси с историком и продюсером Дмитрием Споровым
22 ноября 202490364 В разлуке
В разлукеТри дневника почти за три военных года. Все три автора несколько раз пересекали за это время границу РФ, погружаясь и снова выныривая в принципиально разных внутренних и внешних пространствах
14 октября 202491648 В разлуке
В разлукеМария Карпенко поговорила с экономическим журналистом Денисом Касянчуком, человеком, для которого возвращение в Россию из эмиграции больше не обсуждается
20 августа 202495074 В разлуке
В разлукеСоциолог Анна Лемиаль поговорила с поэтом Павлом Арсеньевым о поломках в коммуникации между «уехавшими» и «оставшимися», о кризисе речи и о том, зачем людям нужно слово «релокация»
9 августа 202494781 В разлуке
В разлукеБыть в России? Жить в эмиграции? Журналист Владимир Шведов нашел для себя третий путь
15 июля 202496084 В разлуке
В разлукеКак возник конфликт между «уехавшими» и «оставшимися», на какой основе он стоит и как работают «бурлящие ритуалы» соцсетей. Разговор Дмитрия Безуглова с социологом, приглашенным исследователем Манчестерского университета Алексеем Титковым
6 июля 202496762 В разлуке
В разлукеФилософ, не покидавшая Россию с начала войны, поделилась с редакцией своим дневником за эти годы. На условиях анонимности
18 июня 2024103443 В разлуке
В разлукеПроект Кольты «В разлуке» проводит эксперимент и предлагает публично поговорить друг с другом «уехавшим» и «оставшимся». Первый диалог — кинокритика Антона Долина и сценариста, руководителя «Театра.doc» Александра Родионова
7 июня 2024101448 В разлуке
В разлукеИван Давыдов пишет письмо другу в эмиграции, с которым ждет встречи, хотя на нее не надеется. Начало нового проекта Кольты «В разлуке»
21 мая 202483239 Colta Specials
Colta Specials Colta Specials
Colta Specials